2023年5月1日 / 最終更新日時 : 2023年8月9日 戎崎 俊一 天体物理学 宇宙における航跡場加速 宇宙から10^20eVを超えるエネルギーを持った超高エネルギー荷電粒子(宇宙線)がやってきている。これは、人工加速器をはるかに超える天然加速器が宇宙には実在していることを意味している。その実体は長年謎のままだったが、最近 […]
2023年4月4日 / 最終更新日時 : 2023年5月2日 戎崎 俊一 その他 アドレス変更のお知らせ 戎崎の科学は一つは、https://science.gakuji-tosho.jpに移動しました。今後はこちらに投稿します。
2023年3月21日 / 最終更新日時 : 2023年8月9日 戎崎 俊一 気候変動 海上低層雲による気候変動緩衝 戎崎(戎崎2023)は、地球の気候が海上低層雲の雲アルベド効果による緩衝により強く安定化されていることを明らかにした。海上低層雲は、海水面温度が低い大陸西岸沖の海洋上にできる表面境界層(雲冠表面境界層)の上部を覆って広がっている。低層雲の存在は、その可視光に対する高いアルベドと、赤外線領域における強い放射冷却で、地球の熱収支を冷却側に強く傾ける。左図に示すように、温室効果ガスの濃度の増加に伴う温暖化は、対流圏・成層圏界面に始まって次第に地上(海上)に波及するが、その過程で低層雲の雲頂に存在する気温逆…
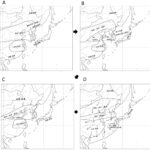
2022年10月18日 / 最終更新日時 : 2023年8月9日 戎崎 俊一 気候変動 日本への水田稲作の伝搬: 環東シナ海文化圏仮説 日本人は、温暖湿潤な日本列島に居住し、代々水田で稲を作って暮らしてきた。その影響は、日本人の生活と文化に深く根差している。水田稲作自身は約10000年前ごろ、中国の長江流域で始まった。従来,水田稲作の伝搬経路に関しては、長江流域で始まった水田稲作が北上して山東半島まで到達し、その後遼東半島,韓半島を経由して日本に到達したという仮説(宮川2017;以後宮川説と呼ぶ)が有力とされてきた。しかし、1) 弥生時代の開始時期が500年早まった、2) 4.2kyrイベントによる寒冷化で水田稲作の北限が南下した、…
2022年10月13日 / 最終更新日時 : 2023年8月9日 戎崎 俊一 地震と津波防災 水中核爆発による津波について ロシアが開発中の核魚雷ポセイドン(約5Mt)による津波のことが巷で話題になっているようなので、定量的に評価してみた。水中核爆発の場合、発生したエネルギーは大部分水の気化に使われる。水が蒸発で失われる半径Rは、R=(3E/4πU)^(1/3)=78 [m](E/TNTMt)^(1/3)と評価できる。ここで、E は爆発エネルギー、U=2.5×10^9 J/m^3は、1立方メートルの水を蒸発するのに必要な気化エネルギーである。このサイズ泡ができ、それが上空に抜けると、それを埋め合わせるように周りから水が流…
2022年3月27日 / 最終更新日時 : 2023年4月29日 戎崎 俊一 時事問題 坂本龍一&吉永小百合が反戦訴え「みんなで絶対にダメだと言ってあらがえばよい状況になる」 吉永小百合さんが出演し、大ヒットした映画「キューポラのある街」(1962年製作)は、最後の方で朝鮮民主主義人民共和国(北朝鮮)を「地上の楽園」と賛美しつつ、彼女が扮する主人公の友人の朝鮮人家族が船に乗って北帰するシーンで終わっている。すぐに続編が作られ、日本に残った上記朝鮮人家族の日本人妻を北帰するよう吉永さんが扮する主人公が説得するシーンもあるという(戎崎は見ていない)。在日朝鮮人の北朝鮮への帰還事業は、1989年まで続いており、家族とともに北朝鮮に渡った日本国籍者は数千人に登るという。その中には…
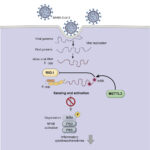
2022年2月8日 / 最終更新日時 : 2023年4月6日 戎崎 俊一 新型コロナ感染症 SARS-CoV-2ウィルスのRNA修飾が宿主細胞の免疫反応を抑制している Li et al. (2021)によると、SARS-CoV-2ウィルスのゲノムは、RNA3’末端領域が強くN6-methylation of Adenosine (m6A)修飾を受けており、これによってウィルス感染による宿主細胞の免疫反応を回避している。彼らは、宿主細胞のm6Aメチル基転移酵素であるMETLL3を減少させると、SARS-CoV-2ウィルスのRNAのm6A修飾が減少し、RIG-Iタンパク分子のRNA分子への結合が増加し、その下流の免疫回路と炎症反応遺伝子の発現を活発化させることを見出し…